
COVID19: Bahaya Varian Delta dan Bagaimana ia Menakluki Dunia
COVID19: Bahaya Varian Delta dan Bagaimana ia Menakluki Dunia
Oleh: Profesor Madya Dr. Azzmer Azzar Bin Abdul Hamid & Ts Dr. Mohd Hamzah Bin Mohd Nasir
Jabatan Bioteknologi, Kulliyyah Sains, Universiti Islam Antarabangsa Malaysia
Hampir dua tahun lamanya kita bergelut dengan pandemik COVID-19 yang berpunca dari coronavirus bernama SARS-CoV-2. Secara kronologi, virus ini dipercayai berasal dari sebuah pasar di Wuhan, China. Ia kemudiannya merebak ke seluruh China dan kemudiannya ke seluruh dunia. Akibat interaksi virus dengan persekitaran, termasuk populasi manusia serata dunia telah menyebabkan berlakunya mutasi kepada genom virus untuk membolehkan ia semakin serasi dengan manusia. Dari varian asal Wuhan kepada varian-varian baharu yang diberi nama dari Alpha, Beta, Gamma dan yang terkini dan paling ditakuti adalah varian Delta.
Mengapa varian Delta lebih bahaya? Untuk menjawab persoalan ini, kita perlu memahami biologi virus serta mekanisma jangkitannya terlebih dahulu. Coronavirus ini adalah virus yang membawa jujukan genom dalam format kimia mRNA. Rantaian genom tersebut dibaluti oleh struktur yang terdiri dari protein envelop (E-Protein), protein membrane (M-Protein) dan protein deduri (Spike protein) (Rajah 1) .Secara asasnya, virus ini berbentuk sfera dan mempunyai struktur deduri yang terbina dari protein yang dinamakan ‘Spike’. Protein ‘Spike’ inilah menjadi perhatian saintis dunia kerana peranannya yang signifikan dalam proses jangkitan sel perumah.
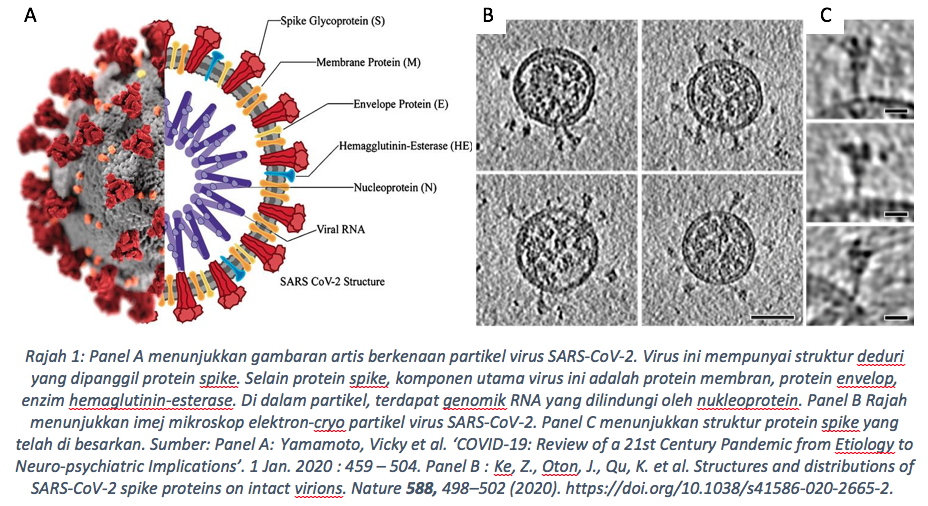
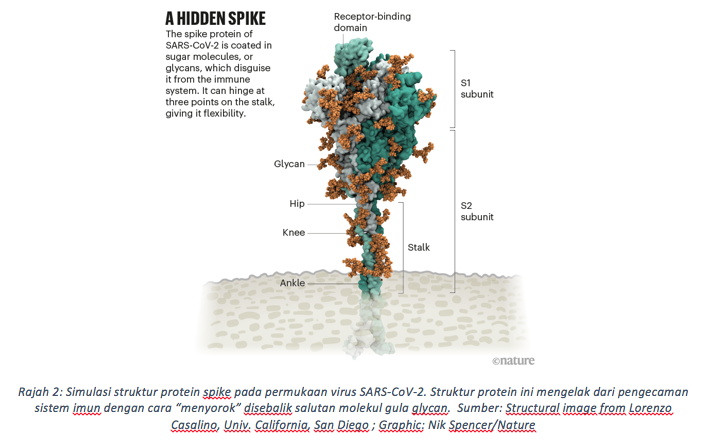
Pada 22 Mac 2020, Professor Rommie Ammaro , seorang ahli pengkomputeran kimia biofizik di Universiti Carlifornia telah mejalankan simulasi atomik melalui maklumat protein Spike yang diperoleh dari jujukan genom virus tersebut (Casalino, Gaieb, Goldsmith et al. 2020, Muratov, Amaro, Andrade et al. 2021). Dari kajian tersebut, beliau mendapati bahawa protein ‘Spike’ ini kelihatan seperti bersembunyi di sebalik molekul gula dari kelas ‘glycan’. Namun, ada satu bahagian pada protein ‘Spike’ tersebut yang tidak diselaputi oleh gula glycan. Hasil perkongsiannya di twitter telah mendapat reaksi seorang ahli biologi struktur dari Universiti Texas, Professor Jason McLellan. Beliau memaklumkan bahawa bahagian protein ‘Spike’ yang terdedah tanpa gula itu adalah domain lekatan reseptor (RBD; receptor binding domain). RBD inilah menjadi struktur terpenting dalam proses ‘berjabat tangan’ antara virus SARS-CoV-2 dan sel manusia yang mempunyai reseptor ACE2 (Angiotensin Converting Enzyme 2). Tidak dinafikan, pengecaman dan interaksi antara RBD pada protein ‘Spike’ dan reseptor ‘ACE2’ pada sel perumah adalah kunci utama proses jangkitan sel, dan mutasi membina pada struktur ini telah meningkatkan kadar transmisi virus seperti yang dilihat pada varian Delta. Varian yang kini sedang mendominasi jangkitan COVID-19 di Malaysia dan seluruh dunia.
Sejak dari awal tercetusnya pandemik COVID-19 ini, saintis dari pelbagai disiplin ilmu telah bertungkus lumus untuk memahami mekanisma jangkitan yang digunakan oleh virus ‘licik’ ini. Majoriti saintis ini berpendapat bahawa dengan memahami mekanisma jangkitan virus ke dalam sel perumah dapat membantu dalam usaha mencari penawar kepada penyakit berjangkit COVID-19 ini. Dari idea inilah para saintis mula menjalankan kajian penggunaan semula ubatan berlainan tujuan (drug repurposing) dalam usaha mengurangkan impak COVID-19 terhadap tubuh.
Bagaimana virus SARS-CoV-2 menjangkiti sel manusia?
Hasil pengetahuan dan kajian mengenai virus SARS-CoV-2 ini telah membawa kepada terbentuknya mekanisma asas jangkitan virus ini pada sel manusia. Rajah 3 menunjukkan ringkasan mekanisma ini.

Berduri untuk menjangkiti
Bermula dari protein Spike. Setiap partikel SARS-CoV-2 mempunyai permukaan luar yang ditunjangi oleh antara 24 dan 40 protein Spike dan bersedia untuk menjangkiti sel perumah. Berbeza dengan virus lain seperti influenza yang mempunyai protein perlekatan permukaan yang tegar, protein ‘Spike’ ini lebih fleksibel (Turoňová, Sikora, Schürmann et al. 2020). Sifat mudah lentur ini membolehkan ianya lebih bebas bergerak mencari sudut sempurna bagi perlekatan yang lebih kuat dan rapat pada sel perumah.
Pada awal kemunculan pandemik, para saintis telah mengesahkan bahawa RBD pada struktur protein ‘Spike’ virus SARS-CoV-2 merupakan lokasi lekatan dengan reseptor ACE2 pada sel yang kebanyakannya berada di saluran pernafasan dan paru-paru manusia. Reseptor yang sama juga digunakan oleh SARS-CoV, virus yang menyebabkan sindrom pernafasan akut teruk (SARS; Severe Acute Respiratory Syndrome). Namun berbanding SARS-CoV dan MERS, RBD pada SARS-CoV-2 melekat antara 2 hingga 4 kali ganda lebih kuat pada reseptor ACE2 akibat mutasi dan penyesuaian yang menstabilkankan interaksi ini (Nguyen, Lan, Thai et al. 2020, Shang, Ye, Shi et al. 2020). Varian-varian virus SARS-CoV-2 lebih cenderung untuk bermutasi pada sub unit S1 pada protein ‘Spike’ yang menjadikan interaksinya bersama reseptor ACE2 lebih teguh dari sebelumnya. Corak mutasi yang sama juga kelihatan pada varian Delta yang kini mula mendominasi jangkitan di seluruh dunia.
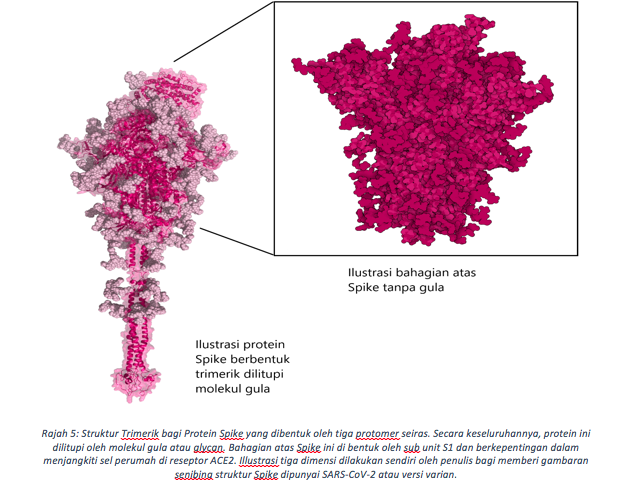
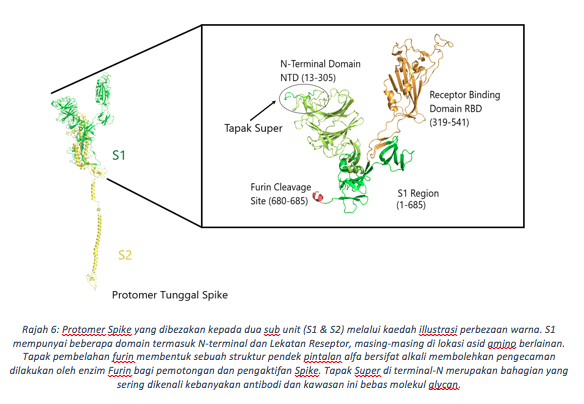
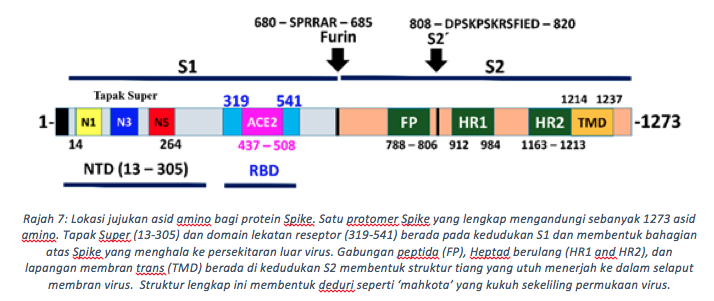
Bagi memahami fungsi Spike ini, kita perlu memahami seni bina strukturnya terlebih dahulu. Protein Spike berbentuk struktur trimerik terhasil dari tiga protomer seiras (Rajah 5). Bagi setiap protomer pada genomik RNA virus ini, sub-unit S1 dan S2 menjadikan ia struktur yang kukuh di atas permukaan virus. S1 mengandungi beberapa lapangan (domain) termasuk N-Terminal dan domain lekatan reseptor (RBD) (Rajah 6). Permukaan RBD dibentuk oleh jujukan asid amino dari susunan 319 hingga 541, dengan sebanyak 17 daripada unit ini berhubung terus dengan ACE2 di permukaan sel manusia (Rajah 7). Tiga gegelung (loops) (N1,N3 dan N5) dalam domain N-terminal (NTD, 13-305 asid amino) ditakrifkan sebagai Tapak Super, iaitu kawasan yang dikenalpasti oleh antibodi yang anti kepada bahagian NTD. Seperti RBD, Tapak super ini merupakan bahagian bebas dari molekul glycan kebiasaanya melitupi hampir setiap permukaan Spike. Tapak pembelahan furin yg unik pula memisahkan S1 daripada terminal-C, domain gabungan membran (FP). Bagi koronavirus biasa, SARS-CoV hanya mengandungi satu asid amino iaitu Arginin di tapak pembelahan ini, manakala koronavirus baru mempunyai penambahan empat (4) asid amino (681-PRRA-684) di hulu (upstream) R685 (Winger & Caspari, 2021). Penghapusan keempat-empat residu ini melalui eksperimentasi telah mengurangkan replikasi virus di barisan sel pernafasan manusia. Oleh itu, penambahan asid amino menggalakkan penyebaran virus di kalangan perumah, sejajar dengan keperluannya dalam penyebaran, dibuktikan oleh ujikaji model haiwan.
Koronavirus baru mempunyai tapak pembelahan furin (680–685aa) yang sudah dibelah seawal zarah virus terbentuk di jasad golgi perumah. Pembelahan Furin melonggarkan struktur trimer tertutup, dengan itu mencetuskan peralihan ke fasa prefusi separa-stabil di mana subunit S1 mengalami gerakan fleksibel. Ini membenarkan domain RBD mengawal atur keadaan tertutup dan terbuka sementara S1 tetap terikat dengan S2. Pembelahan tapak S2’ (808-820) ketika berlakunya pengikatan kepada ACE2, memisahkan subunit S1 dari S2 dengan efisien. Cebisan S2 yang dihasilkan kemudian mengalami perubahan konformasi yang ketara di mana dua heptad berulang (HR1, HR2) membentuk domain gabungan enam helix, dan peptida gabung (FP) (788-806aa) membolehkan virus bersatu dengan sel sasaran di plasma membran. Maka berlakulah penggabungan antara virus dan sel manusia.
Kemasukan virus ke sel perumah.
Terdapat dua mekanisma asas kemasukan virus kedalam sel manusia telah dikenalpasti. Namun pada varian Delta, mekanisma utama jangkitan virus ke dalam sel perumah bergantung pada enzim kelas protease TMPRSS2 yang terdapat pada permukaan sel perumah, terutamanya pada sel epitelium saluran pernafasan. Kelebihan menggunakan TMPRSS2 secara efisien oleh SARS-CoV-2 ini menjadikan mekanisma jangkitan pada virus penyebab COVID-19 ini berbeza berbanding daripada SARS dan MERS terdahulu.
Membran virus dan sel perumah mula bersatu apabila enzim TMPRSS2 pada sel perumah memangkas protein ‘Spike’ pada perumah. Akibatnya, asid amino yang bersifat hidrofobik pada protein ‘Spike’ akan terdedah dan mula mencari membran terhampir yang juga bersifat hidrofobik untuk bergabung (Rajah 8). Tindak balas spontan ini menyebabkan tergabungnya membran pada virus dan sel perumah. Penggabungan membran ini membolehkan virus memasukkan jujukan genom mRNA ke dalam sitoplasma sel perumah.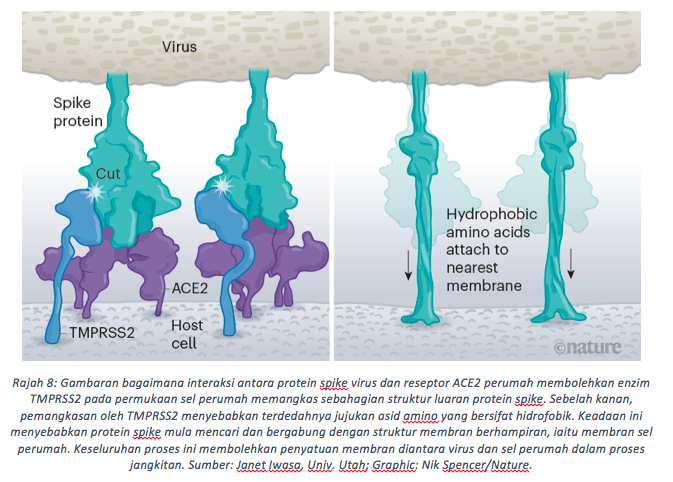
Rampasan kuasa ke atas perumah
Proses selepas penggabungan membran adalah lebih menarik dan menakutkan. Apabila genom RNA virus dipindahkan ke dalam sel perumah, ribosom yang terdapat pada sitoplasma mula mentakrif dua bahagian kod mRNA tersebut kepada rantaian asid amino yang kemudiannya dipotong menjadi 16 protein berbeza, termasuk protein yang diperlukan untuk penggandaan RNA virus. Daripada protein yang dihasilkan ini, lebih banyak RNA terhasil dan digunakan untu pengkodan 26 protein, termasuk protein stuktur (Protein spike, protein Envelop dan protein Membran sebagai contoh) yang diperlukan untuk penghasilan partikel virus yang baharu.
Untuk mengelakkan persaingan penggunaan ribosom untuk penggandaan komponen sel perumah, protein Nsp1 yang berasal dari virus menghapuskan mRNA lain yang tidak mempunyai tag virus (Finkel, Gluck, Nachshon et al. 2021). Ini bermakna virus mampu merampas ribosom sel perumah untuk hanya menghasilkan protein virus sahaja.
Bertambah teruk, protein Nsp1 ini juga menutup laluan keluar mRNA perumah dari nukleus (Zhang, Miorin, Makio et al. 2021). Ini menyebabkan segala tindak balas korektif dari sel perumah tehadap jangkitan disenyapkan dan dilumpuhkan. Akibatnya, sel tidak dapat menghasilkan protein-protein kendiri bagi tujuan komunikasi antara sel termasuk interferon yang digunakan antaranya untuk memberitahu sistem imun akan jangkitan virus di dalam sel.
Disamping penghasilan protein struktur virus, genom RNA virus juga digandakan di dalam sel perumah. Ini membolehkan lebih partikel virus baharu dihasilkan dengan cekap dan pantas. Keadaan ini membantu virus menghasilkan komponen yang diperlukan dengan banyak dan berkesan.
Pengubahsuaian sel perumah
Apabila virus telah berjaya merampas sistem translasi protein pada sel perumah, tanpa berlengah, virus memulakan proses menjadikan sel perumah sebagai kilang penghasilan partikel virus yang baharu. Protein ‘Spike’ yang dihasilkan tadi akan dieksport ke permukaan sel dan mengaktifkan liang ion-kalsium (Calcium-ion channel). Fenomena ini menyebabkan sel yang dijangkiti ini mula bergabung dengan sel bersebelahan yang mempunyai ACE2, dan membentuk gagasan sel yang besar dan mempunyai nukleus yang banyak (Braga, Ali, Secco et al. 2021).
Dalam pada masa yang sama, protein virus dan mRNA virus yang dihasilkan tadi mula bergabung didalam organel pemprosesan protein pada sel perumah, iaitu retikulum endoplasmik dan jasad Golgi. Disinilah partikel baharu virus mula dikumpulkan dan di’pasang’. Genom RNA virus yang terhasil dari proses replikasi RNA mula berpasangan dan distabilkan oleh molekul protein nukleokapsid, antara molekul yang digunakan sebagai petanda pengesanan didalam RTK-antigen SARS-COV-2. Struktur RNA-nukleokapsid ini akan menuju kearah jasad Golgi yang mempunyai protein M dan protein S, serta protein struktur yang lain. Penggabungan kesemua komponen ini membentuk satu unit partikel virus yang baharu. Walaubagaimanapun, dalam konfigurasi asas ini virus masih dikatakan sebagai tidak aktif dan tidak mampu untuk menjangkiti sel lain. Satu modifikasi ringkas diperlukan pada struktur protein’Spike’ untuk menjadikannya aktif .
Suntingan terakhir.
Dalam perjalanan keluar dari sel perumah, satu lagi suntingan terakhir diperlukan. Partikel virus yang terhasil di jasad Golgi membawa protein ‘Spike’ dalam konfigurasi tidak aktif. Untuk mengaktifkannya, bahagian S1 dan S2 protein ‘Spike’ perlu diputuskan pada kedudukan pemisah yang dipanggil sebagai tapak pembelahan furin (Hoffmann, Kleine-Weber and Pöhlmann 2020, Mykytyn, Breugem, Riesebosch et al. 2021)
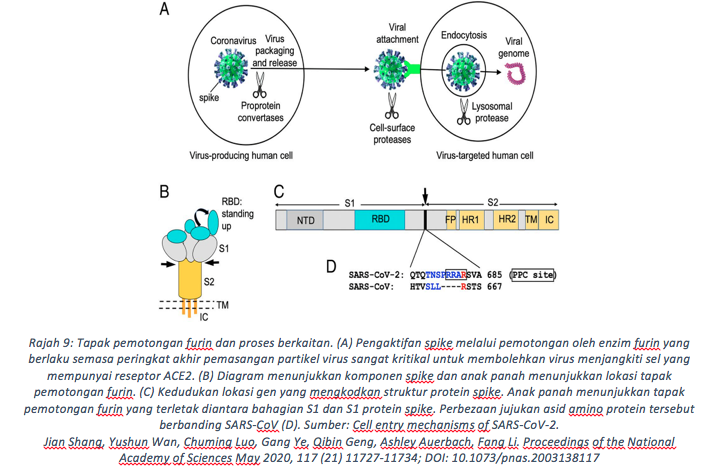
Suntingan ini berlaku antara ketika proses ‘pemasangan’ virus di jasad Golgi dan pelepasan virus keluar dari sel perumah kerana enzim furin ini boleh didapati di dalam vesikel yang terhasil di organel tersebut. Dengan pemotongan ikatan peptida di tapak pembelahan furin, ianya melonggarkan sedikit struktur protein ‘Spike’ dan membolehkan RBD mengesan ACE2 dan memulakan interaksi yang melibakan pemprosesan oleh TMPRSS2 seperti yang dimaklumkan dia awal artikel ini.
Sehingga kini, varian Alfa dan Delta telah diketahui mempunyai mutasi satu asid amino pada kedudukan jujukan protein 681 di tapak pemotongan furin. Varian alfa membawa mutasi P681H dimana asid amino proline ditukar kepada histidin. Varian delta pula bermutasi di P681R yang mengubah asid amino proline kepada arginin. Kedua-dua mutasi ini menyebabkan rantaian protein pada tapak pemotongan furin bersifat lebih beralkali dan membolehkan pemotongan yang lebih efektif oleh enzim furin.
Pemotongan furin yang lebih efisien meningkatkan kesiapsediaan virus untuk menjangkiti sel perumah. Didapati, jumlah protein ‘Spike’ yang teraktif pada SARS-CoV-2 adalah lebih 40% berbanding pada SARS-CoV. Jumlah ini meningkat sebanyak 50% pada varian Alfa manakala melebihi 75% bagi varian Delta. Hubungan ini membina hipotesis dimana perkaitan jumlah spike teraktif melalui pemotongan furin diantara struktur S1 dan S2 berkadaran dengan peningkatan kebolehjangkitan SARS-CoV-2 pada sel perumah.
Corak mutasi dan kaitan kebolehjangkitan
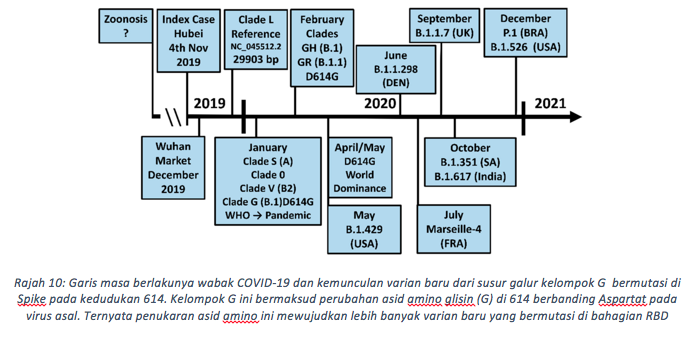
Pada mulanya, kemunculan varian baru tercetus akibat peningkatan mutasi global, D614G (Rajah 10). Ia pertama kali dikesan bermutasi secara bebas pada Januari 2020 di China dan Jerman, dan berjaya mengatasi virus Wuhan menjelang April / Mei 2020. Mutasi ini mentakrifkan kluster baru iaitu kluster G (G, GH, GR, GV) yang menggantikan kluster L. Kelompok GR menguasai Afrika (41.1%), Asia (52.7%), Oceania (74.8%), dan Amerika Selatan (66.8%), sementara GH mendominasi Amerika Utara (59.0%). Kedua-dua kelompok bagi GR (35.5%) dan GV (34.6%) menjadi varian bersama yang berjaya menjangkiti manusia secara kodominan di Eropah (Winger and Caspari 2021). Di Malaysia, kelompok G ini sangat berjaya mengembangkan keturunan virus baru termasuk varian alpha, beta, dan delta. Terkini, varian delta sedang menguasai jangkitan di Malaysia sebanyak 77%, diikuti beta 17% dan varian lain iaitu 6% berdasarkan jumlah sampel penjujukan diperolehi dalam tempoh dua bulan bermula June. Melalui penjujukan genom yang dibuat, secara keseluruhan sebanyak 17,486 genom delta telah dikesan dalam pesakit COVID-19 di dunia. Varian delta ini sedang giat menjangkiti manusia dan akan mendominasi dunia dengan kadar pantas, tidak pernah dilihat pada koronavirus sebelumnya.
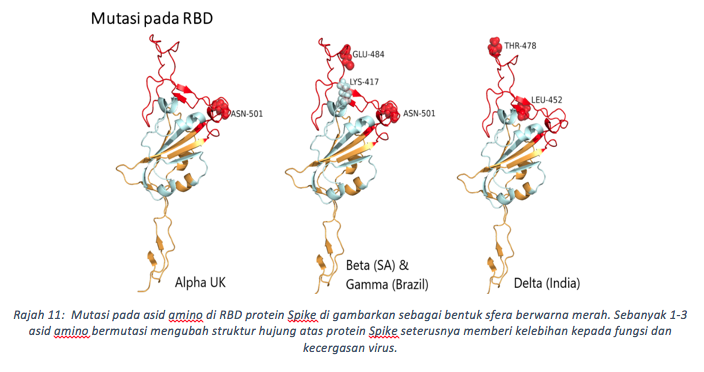
Penggantian Aspartat-614 yang bercas negatif kepada asid amino glisin yang lebih kecil dan tidak bercas (D614G) menjejaskan ikatan dengan lisin-854 (K854) dan T859, keduanya di kedudukan protomer bersebelahan. Perubahan struktur secara keseluruhan tertumpu kepada gegelung 620-640 asid amino yang berdekatan dengan tapak pembelahan furin. Gelung ini lebih stabil untuk G614 Spike, kerana ia sesuai dengan bukaan yang lebih besar disebabkan glisin yang lebih kecil. Ini tidak dapat ditemukan dalam virus wuhan D614. Walaupun terdapat perubahan kecil, impak kepada kecergasan virus adalah sangat tinggi. Sementara virus wuhan secara purata mempunyai satu bukaan RBD setiap masa, G614 pula mempunyai dua atau ketiga-tiga RBD dalam sifat terbuka dan bersedia menjangkiti ACE2. Mutasi D614G juga mengurangkan pembebasan S1 pramatang, dan meningkatkan kestabilan haba bagi Spike. Gabungan ciri ini meningkatkan jumlah virus di saluran pernafasan hidung dan trakea. Ini menjelaskan mengapa G614 virus sangat cergas berbanding virus D614, terbukti melalui ujikaji persaingan menggunakan model haiwan. Setelah inokulasi haiwan diuji dengan nisbah sama antara virus (D614 & G614), lebih 90% virus yang disebarkan membawa Spike G614. Oleh itu, transmisi virus membawa G614 sangat mudah berlaku dan menjangkiti sel perumah. Varian yang diberi perhatian (VOC) kini sebenarnya muncul dari kelompok G (B.1) D614G, antaranya adalah Alpha (B.1.117), Beta (B.1.351), Gamma (P.1) dan terkini Delta (B.1.617). Kesemua varian ini mempunyai mutasi di sepanjang genom virus terutama pada bahagian Spike, iaitu di kedudukan RBD dan bukan RBD (Rajah 11). Apa yang menarik, kebanyakan lokasi mutasi mempunyai beberapa kesamaan jika dibandingkan dengan varian-varian yang sedang berlegar di dunia (rajah 12). Pertukaran asid amino ini pada RBD diberi perhatian penuh oleh penyelidik kerana berkemungkinan secara langsung meningkatkan pelekatan reseptor. Bagi varian delta, dua asid amino di RBD telah berubah (kedudukan L452R dan T478K), dan satu asid amino (681) berdekatan dengan tapak pembelahan furin ditukar kepada residu beralkali iaitu Arginin. Kesannya seperti yang dapat kita kaitkan, kadar kebolehjangkitan bagi varian delta menunjukkan tren paling tinggi berbanding varian lain yang pernah ditemui.

Varian delta lebih mudah disebarkan berada di landasan tepat untuk menjadi dominan di seluruh dunia. Pertama kali dilihat di India dan sekarang di sekurang-kurangnya 135 negara, penyebarannya yang meluas telah menyebabkan sekatan baru di seluruh dunia. Kebimbangan tentang varian delta berlaku di pelusuk dunia, begitu juga dinyatakan oleh Tedros Adhanom Ghebreyesus, ketua pengarah Organisasi Kesihatan Sedunia (WHO). Ketika ini, delta dicatatkan sebagai varian perhatian kedua paling dominan selepas alpha, kira-kira sebanyak 80,000 kes. Dominasi varian delta di UK dengan pantas menunjukkan kadar vaksinasi yang tinggi tidak menjadi halangan besar buat virus ini. Sehingga Julai, varian ini menyumbang kepada 95 peratus dari semua kes baru di UK. Pusat Pencegahan dan Pengawalan Penyakit di Eropah membuat unjuran, pada akhir bulan Ogos, delta akan membawa sebanyak 90 peratus kes COVID-19 di Kesatuan Eropah. Sekatan perjalanan juga dilaksanakan di Greater Sydney, Australia, berikutan lebih dari 100 kes varian baru. Manakala Israel telah memperkenalkan semula pemakaian topeng muka hanya setelah 10 hari menamatkannya, berikutan susulan kes import. Di Asia, kes meningkat dengan cepat di Bangladesh, Iran, Iraq, Jepun, Kazakhstan, Malaysia, Myanmar, Pakistan, Selatan Korea, Thailand dan Vietnam. Antara yang terkesan, negara paling tinggi rakyatnya diimunisasi iaitu Jepun, dengan 35% penduduk diberi vaksin dos lengkap. Jiran negara kita, Indonesia, telah menvaksinasi sepenuhnya 7.9% warganya, namun begitu sebanyak lebih seribu kematian harian direkodkan sejak ogos. Dari segi per kapita, ini adalah sama dengan kadar kematian puncak yang dilaporkan oleh India di pertengahan Mei — 3.32 kematian per juta orang setiap hari (Dyer 2021).
Penyelidik berpendapat varian delta begitu pantas menjangkiti kerana berjaya menghasilkan virus baru dengan kuantiti yang banyak berbanding SARS-CoV-2 versi asal. Anggaran penyebaran varian delta adalah lebih dari dua kali ganda berbanding virus Wuhan. Ahli epidemiologi, Jing Lu dari Pusat Kawalan dan Pencegahan Penyakit Guangzhou, China, dan rakannya telah mengesan 62 pesakit COVID-19 yang dikuarantin dijangkiti varian Delta. Pasukan ini menguji ‘jumlah virus’ pesakit dengan mengukur ketumpatan virus dalam badan – setiap hari sepanjang jangkitan untuk melihat bagaimana ia berubah mengikut masa. Proses penyelidikan ini juga melibatkan perbandingan pesakit dijangkiti virus asal, SARS-CoV-2. Dalam laporan mereka, variant delta pertama kali dikesan empat hari selepas pendedahan, berbanding dengan purata enam hari oleh orang yang dijangkiti virus asal, menunjukkan bahawa delta mereplikasi lebih pantas. Individu yang dijangkiti delta juga mempunyai jumlah virus hingga 1260 kali lebih tinggi (Reardon 2021). Gabungan kuantiti virus dan tempoh inkubasi pendek menjelaskan bagaimana varian delta tersebar dengan begitu pantas dan mudah merebak. Jumlah virus yang tinggi pada saluran pernafasan menyokong penyebaran super kepada lebih ramai orang, dan apablia dijangkiti, mereka bakal menyebarkan virus seawal tempoh jangkitan. Inkubasi pendek menjadikan ia lebih sukar dibendung, menjejaskan sistem saringan kerajaan. Ada beberapa persoalan mengenai varian delta masih belum terjawab. Ia masih tidak jelas, misalnya, adakah varian ini lebih cenderung menyebabkan penyakit yang teruk berbanding virus SARS-CoV asal, dan bagaimana varian ini menghindari sistem ketahanan badan kita. Maklumat ini akan diperolehi apabila para penyelidik mengkaji secara mendalam dengan melibatkan populasi manusia yang lebih besar. Sehingga hari ini, varian delta ini benar-benar mengejutkan dunia dan secara keseluruhan, ia sangat sukar dihentikan.
Kesimpulan
Melalui artikel ini, jelas menjukkan bahawa kejayaan mutasi pada varian Delta telah menjadikan COVID-19 ini lebih mudah merebak, menjangkiti manusia serta menyebabkan kesan kesihatan awam yang teruk. Walaupun usaha vaksinasi sedang giat dilaksanakan oleh agensi yang berkaitan, usaha membendung jangkitan yang semakin ganas ini juga terletak pada setiap individu itu sendiri. Pematuhan rapi terhadap SOP yang disarankan oleh Kementerian Kesihatan Malaysia adalah antara langkah yang sepatutnya dipandang serius oleh setiap lapisan masyarakat. Elakkan diri dari dijangkiti dan elakkan diri dari menjangkiti.
Nota: Sebahagian artikel ini diolah dari: ‘How the coronavirus infects cells — and why Delta is so dangerous’, Megan Scudellari (28 Julai 2021)
RUJUKAN
Braga, Luca, Hashim Ali, Ilaria Secco, Elena Chiavacci, Guilherme Neves, Daniel Goldhill, Rebecca Penn, Jose M Jimenez-Guardeño, Ana M Ortega-Prieto and Rossana Bussani. 2021. “Drugs That Inhibit Tmem16 Proteins Block Sars-Cov-2 Spike-Induced Syncytia.” Nature 594(7861):88-93.
Casalino, Lorenzo, Zied Gaieb, Jory A Goldsmith, Christy K Hjorth, Abigail C Dommer, Aoife M Harbison, Carl A Fogarty, Emilia P Barros, Bryn C Taylor and Jason S McLellan. 2020. “Beyond Shielding: The Roles of Glycans in the Sars-Cov-2 Spike Protein.” ACS Central Science 6(10):1722-34.
Finkel, Yaara, Avi Gluck, Aharon Nachshon, Roni Winkler, Tal Fisher, Batsheva Rozman, Orel Mizrahi, Yoav Lubelsky, Binyamin Zuckerman and Boris Slobodin. 2021. “Sars-Cov-2 Uses a Multipronged Strategy to Impede Host Protein Synthesis.” Nature 594(7862):240-45.
Hoffmann, Markus, Hannah Kleine-Weber and Stefan Pöhlmann. 2020. “A Multibasic Cleavage Site in the Spike Protein of Sars-Cov-2 Is Essential for Infection of Human Lung Cells.” Molecular cell 78(4):779-84. e5.
Muratov, Eugene N, Rommie Amaro, Carolina H Andrade, Nathan Brown, Sean Ekins, Denis Fourches, Olexandr Isayev, Dima Kozakov, José L Medina-Franco and Kenneth M Merz. 2021. “A Critical Overview of Computational Approaches Employed for Covid-19 Drug Discovery.” Chemical Society Reviews.
Mykytyn, Anna Z, Tim I Breugem, Samra Riesebosch, Debby Schipper, Petra B van den Doel, Robbert J Rottier, Mart M Lamers and Bart L Haagmans. 2021. “Sars-Cov-2 Entry into Human Airway Organoids Is Serine Protease-Mediated and Facilitated by the Multibasic Cleavage Site.” Elife 10:e64508.
Nguyen, Hoang Linh, Pham Dang Lan, Nguyen Quoc Thai, Daniel A Nissley, Edward P O’Brien and Mai Suan Li. 2020. “Does Sars-Cov-2 Bind to Human Ace2 More Strongly Than Does Sars-Cov?”. The Journal of Physical Chemistry B 124(34):7336-47.
Shang, Jian, Gang Ye, Ke Shi, Yushun Wan, Chuming Luo, Hideki Aihara, Qibin Geng, Ashley Auerbach and Fang Li. 2020. “Structural Basis of Receptor Recognition by Sars-Cov-2.” Nature 581(7807):221-24.
Turoňová, Beata, Mateusz Sikora, Christoph Schürmann, Wim JH Hagen, Sonja Welsch, Florian EC Blanc, Sören von Bülow, Michael Gecht, Katrin Bagola and Cindy Hörner. 2020. “In Situ Structural Analysis of Sars-Cov-2 Spike Reveals Flexibility Mediated by Three Hinges.” Science 370(6513):203-08.
Winger, Anna and Thomas Caspari. 2021. “The Spike of Concern—the Novel Variants of Sars-Cov-2.” Viruses 13(6):1002.
Zhang, Ke, Lisa Miorin, Tadashi Makio, Ishmael Dehghan, Shengyan Gao, Yihu Xie, Hualin Zhong, Matthew Esparza, Thomas Kehrer and Anil Kumar. 2021. “Nsp1 Protein of Sars-Cov-2 Disrupts the Mrna Export Machinery to Inhibit Host Gene Expression.” Science Advances 7(6):eabe7386.





{kind=link}